|
|
|
|
|
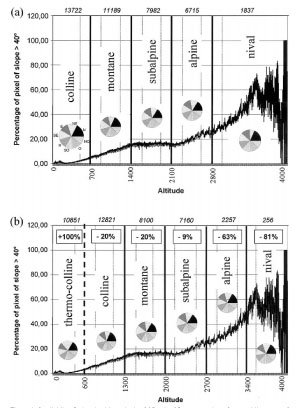
1 - Area shift of vegetation belts in Switzerland (164K) |
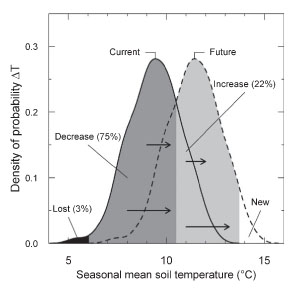
Based on the assumption that vegetation zones will shift corresponding to temperature increase, a 3.3 K annual temperature warming would result in a reduction in the extent of alpine area of Switzerland by 63%, and of nival area by 81% (fig 1). On the other hand, microtopographical differences are quite strong, with surface temperature differences of 8.4±2.5 K (mean±SD) persisting over several hours per day along horizontal transects on bright days (data obtained from six alpine (Alps) and arctic-alpine slopes (Norway, Sweden, Svalbard). Within a given area, only a few percent of microtopographical conditions will be lost (fig. 2) and plants and animals may have to migrate only along short distances to reach suitable microhabitats, even though the abundance of thermally suitable microhabitats will be reduced for many species. So, to which extent will ranges of single species and whole communities change in future? |
2 - The density distribution of current seasonal mean soil temperatures (current) and in a 2 K warmer scenario (future). 'Lost’ indicates the micro-habitat temperatures which disappear in a 2 K warmer scenario within the study area, ‘decrease’ indicates the micro-habitat temperatures that decrease in their abundance, ‘increase’ the ones that increase in abundance and ‘new’ denotes the micro-habitat conditions that are warmer than the current warmest temperatures. The arrows indicate the direction of decreasing and increasing micro-habitat temperatures |
|
An upward range shift of alpine plant species is especially obvious when present-day species lists are compared with historical ones, which has been done for many summits in the Alps but also for other mountains all over the world. In most cases, a strong increase in species richness and abundance has been found (table 1). The vegetation of 150 summits in south-eastern Switzerland has been mapped and compared to historical records. Overall, about 250 plant species have been recorded, with remarkable differences between summits: the richness of the summit flora varied between one and 118 species in the top 10 m of the summit. In conclusion, plant species have clearly advanced to higher elevations, leading to an overall increase in species richness in the summit areas. Many plant species that were already quite common previously have become even more common, e.g. Alpine Bluegrass (Poa alpina) which is now found on 111 summits compared to 59 summits about 100 years ago. Also, some plants from lower altitudes, which had not been recorded previously, became quite numerous. On the other hand, a few plant species declined in abundance for unknown reasons. |
3 - Piz Linard |

|
Table 1 - Plant species present on the summit of Piz Linard (top 30 m) from 1835-2011. Abundances are indicated by colour: light grey, <5 individuals, intermediate grey, <10 individuals, dark grey, >10 individuals (no abundances available for 1864 and 1895). Species present in the top 10 m are indicated with double asterisks (from Wipf 2013). 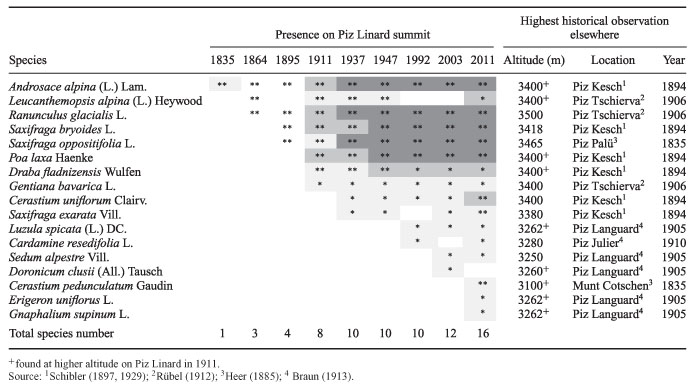
|
|
Many studies compared the actual elevational distribution of plant and animal species with historical data. Migration rates markedly differ between species and sites and range from negative (i.e. downslope migration) to an upslope migration of 4.5 m yr-1. But usually there is an overall upward shift at a rate of several meters per decade up to 2-3 m yr-1. A recent meta-analysis including data from birds, mammals, arthropods, plants, herptiles, fish, and mollusks revealed that species migrated to higher elevations at a median rate of 11 meters per decade, which is somewhat less than the expected range shift if species were able to migrate exactly corresponding to the warming climate. |
|
Factors representing possible limitations for species migration include habitat fragmentation and dispersal limitation, slow soil formation at high altitudes, disturbances e.g. due to thawing of permafrost, as well as a general decline in available area with increasing elevation. Migration can also be hampered by site conditions, where no open space is available for recruitment, or seedling establishment is limited by water availability under increasingly dry conditions. Migration rates considerably differ within and among species, depending on site coditions. For plants, anemochorous species were found to migrate more often and to relatively higher altitudes than zoochorous or autochourous species, and migration was higher on calcareous than on siliceous bedrock. Some late successional species like Carex curvula seem to be quite persistent and do not move at all. Lower range margins shifted upward or downward. For example, the lower range margin of Ranunculus glacialis moved upwards in Scandinavia, probably due to increased competition by other plants. |
4 - Increase in vascular plant species numbers per altitudinal range of 100 m (82K) |
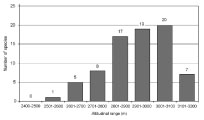
|
Probable causes for species migration As the migration rate has been higher in regions with more pronounced climate warming, and also has accelerated during the last decades parallel to warming, warming most likely is a main driver for these range changes. Yet there are probably other causes that may have contributed to species migration. For example, changes in land use can locally be of great importance (→ 31n). Tree expansion at the treeline often corresponds to abandonment of intensive land use. The increasing number of species at high elevations in Hawaii seems to be more favoured by substrate amelioration and increased traffic accompanied with propagule transport than climate warming. Also, area changes of some insects can be attributed to land use change. |
|
Changes at the community level The community mosaic in the Alpine may shift considerably. At a high alpine site in Italy vegetation changes between 1953 and 2003 (table 2) included an expansion of shrubs between 2200 and 2500 m. Likewise, grassland showed an upward migration, but generally decreased in area. Snowbed vegetation also decreased in area and shifted to higher elevations, whereas bare ground increased in extension. There was an overall vegetation regression above 2600 m, which was probably due to increased disturbances caused by an increase in precipitation and permafrost degradation. Table 2 - Changes (%) in the ecological series of habitat types between 1953 and 2003 for various altitudinal ranges (from Cannone 2007)
|
|
It is often predicted that with increasing temperature the treeline will move upslope. Possible causes for static treelines include that the actual treeline position may not be temperature dependent, but constrained e.g. by edaphic or microclimatic conditions. |
5 - Dovrefjell, Norway (197K) |

|
Treeline advances may also be constrained by large herbivores: Tree ring and photo analyses revealed that treeline advance at a location in northern Sweden was pronounced during relatively cool periods in the second half of the last century and was significantly correlated with periods of low reindeer (Rangifer tarandus) population numbers. Also, other disturbances like avalanches or debris flows can impede tree regeneration, and some sort of closed alpine vegetation has been shown to have a high resilience to the invasion of trees. Proximity to the ocean also has a remarkable influence on the outcome of community development in the face of increasing temperatures, particularly in northern regions. Whereas treeline altitude and tree density increased in more continental areas, in several oceanic areas downward movement of the treeline position in favour of wet heaths, bogs and wetland tundra has been observed. Dehardening and high respiratory loss of the trees due to mild winter temperatures are the probable causes. Irrespective of a potential upward shift, an infilling can be observed at most treeline sites. Also, many trees at the treeline show wider tree-rings in the last decades, which can be attributed to ameliorated growth conditions. In the Urals, Siberian larch (Larix sibirica) shifted not only upwards, but growth form of larch changed from a formerly creeping form to now multi-stemmed upright growth, and single-stemmed trees have commenced to establish, which is the typical growth form for more benign habitats. Age distribution analyses indicate that recruitment in forest ecosystems is mostly episodic rather than gradual close to the tree line. Obviously, several years with favourable conditions are necessary for young trees to establish. Tree development will be impeded by extreme events like drought, severe frost during the growing season or mass outbreaks of leaf-eating insects and pathogens. Thus, the occurrence of severe climatic events is likely to control tree population dynamics to a larger extent than mean temperature increase. |
|
Summit flora in a changing climate | |||
13 August 2018 |
||
| |
||